Функции макрофагов при воспалении

Основная роль в развитии и поддержании хронического воспаления принадлежит системе фагоцитирующих макрофагов (это понятие заменило широко применявшийся ранее, но по существу недостаточно обоснованный термин «ретикулоэндотелиальная система»). Основная клетка этой системы—макрофаг, развившийся из моноцита крови. Моноциты, происходящие из стволовой клетки костного мозга, поступают вначале в периферическую кровь, а из нее в ткани, где под влиянием различных местных стимулов превращаются в макрофаги.
Последние имеют чрезвычайно большое значение в осуществлении адаптивных реакций организма — иммунных, воспалительных и репаративных. Участию в подобных реакциях способствуют такие биологические свойства макрофагов, как способность мигрировать в очаги воспаления, возможность быстрого и стойкого увеличения продукции клеток костным мозгом, активный фагоцитоз чужеродного материала с быстрым расщеплением последнего, активация под действием чужеродных стимулов, секреция ряда биологически активных веществ, способность «обрабатывать» проникший в организм антиген с последующей индукцией иммунного процесса.
Принципиально важно также, что макрофаги являются долгоживущими клетками, способными длительно функционировать в воспаленных тканях. Существенно, что они способны пролиферировать в очагах воспаления; при этом возможна трансформация макрофагов в эпителиоидные и гигантские многоядерные клетки.
Не обладая иммунологической специфичностью (как Т- и В-лимфоциты), макрофаг действует в качестве неспецифической вспомогательной клетки, обладающей уникальной способностью не только захватывать антиген, но и обрабатывать его так, что последующее распознавание этого антигена лимфоцитами значительно облегчается. Этот этап особенно необходим для активации Т-лимфоцитов (для развития иммунных реакций замедленного типа и для продукции антител к тимусзависимым антигенам).
Кроме участия в иммунных реакциях за счет предварительной обработки антигена и его последующего «представления» лимфоцитам, макрофаги осуществляют защитные функции и более непосредственно, уничтожая некоторые микроорганизмы, грибы и клетки опухолей.
Таким образом, при ревматических заболеваниях в клеточных реакциях иммунного воспаления участвуют не только специфически иммунизированные лимфоциты, но и не имеющие иммунологической специфичности моноциты и макрофаги.
Эти клетки привлекаются моноцитарными хемотаксическими веществами, вырабатываемыми в очагах воспаления. К ним относятся С5а, частично денатурированные белки, калликреин, активатор плазминогена, основные белки из лизосом нейтрофилов Т-лимфоциты вырабатывают подобный фактор при контакте ее специфическим антигеном, В-лимфоциты — с иммунными комплексами.
Кроме того, лимфоциты продуцируют также факторы угнетающие миграцию макрофагов (т. е. фиксирующие их в очаге воспаления) и активирующие их функцию. В воспалительных очагах в отличие от нормальных условий наблюдаются митозы макрофагов и таким образом количество этих клеток нарастает также за счет местной пролиферации.
Значение макрофагов в поддержании воспалительного процесса определяется рассматриваемыми ниже противовоспалительными агентами, освобождаемыми из этих клеток.
1. Простагландины.
2. Лизосомные ферменты (в частности, при фагоцитозе комплексов антиген — антитело, причем клетка при их выделении не разрушается).
3. Нейтральные протеазы (активатор плазминогена, коллагеназа, эластаза). В норме их количество ничтожно, но при чужеродной стимуляции (при фагоцитозе) продукция данных ферментов индуцируется и они выделяются в значительных количествах. Продукция нейтральных протеаз угнетается ингибиторами белкового синтеза, в том числе глюкокортикостероидами. Выработка активатора плазминогена и коллагеназы стимулируется также факторами, секретируемыми активированными лимфоцитами.
4. Фосфолипаза Аз, освобождающая из более сложных комплексов арахидоновую кислоту — основной предшественник простагландинов. Активность этого фермента тормозится глюкокортикостероидами.
5. Фактор, стимулирующий освобождение из костей как минеральных солей, так и органической основы костного матрикса. Этот фактор реализует свое влияние на костную ткань за счет прямого воздействия, не требуя присутствия остеокластов.
6. Ряд компонентов комплемента, которые активно синтезируются и выделяются макрофагами: С3, С4, С2 и, по-видимому, также С1 и фактор В, необходимый для альтернативного пути активирования комплемента. Синтез этих компонентов повышается при активировании макрофагов и тормозится ингибиторами белкового синтеза.
7. Интерлейкин-1, который является типичным представителем цитокинов — биологически активных веществ полипептидной природы, вырабатываемых клетками (прежде всего клетками иммунной системы). В зависимости от источников продукции этих веществ (лимфоциты или моноциты) нередко применяются термины «лимфокины» и «монокины». Название «интерлейкин» с соответствующим номером используется для обозначения конкретных цитокинов — особенно тех, которые опосредуют клеточное взаимодействие. Пока не вполне ясно, представляет ли интерлейкин-1, являющийся наиболее важным монокином, одно вещество или семейство полипептидов, обладающих очень близкими свойствами.
К этим свойствам относятся следующие:
- стимуляция В-клеток, ускоряющих их трансформацию в плазматические клетки;
- стимуляция активности фибробластов и синовиоцитов с повышенной выработкой ими простагландинов и коллагеназы;
- пирогенное влияние, реализующееся в развитии лихорадки;
- активирование синтеза в печени острофазовых белков, в частности сывороточного предшественника амилоида (этот эффект, возможно, является опосредованным — благодаря стимуляции выработки интерлейкина-6).
Среди системных эффектов интерлейкина-1, помимо лихорадки, могут быть отмечены также нейтрофилез и протеолиз скелетных мышц.
8. Интерлейкин-6, который также активирует В-клетки, стимулирует гепатоциты к выработке острофазовых белков и обладает свойствами b-интерферона.
9. Колониестимулирующие факторы, способствующие образованию в костном мозге гранулоцитов и моноцитов.
10. Фактор некроза опухолей (ФНО), который не только действительно способен вызывать некроз опухолей, но и играет заметную роль в развитии воспаления. Этот полипептид, состоящий из 157 аминокислот, в раннюю фазу воспалительной реакции способствует прилипанию нейтрофилов к эндотелию и способствует тем самым их проникновению в очаг воспаления. Он служит также мощным сигналом к выработке токсичных кислородных радикалов и является стимулятором В-клеток, фибробластов и эндотелия (2 последних типа клеток при этом вырабатывают колониестимулирующие факторы).
Клинически важно, что ФНО, так же как интерлейкин-1 и интерферон, подавляют активность липопротеинлипазы, которая обеспечивает отложение жира в организме. Именно поэтому при воспалительных заболеваниях часто отмечается выраженное похудание, не соответствующее калорийному питанию и сохранившемуся аппетиту. Отсюда второе название ФНО — кахектин.
Активация макрофагов, проявляющаяся увеличением их размера, большим содержанием ферментов, нарастанием способстности к фагоцитозу и уничтожению микробов и опухолевых клеток, может быть и неспецифичной: за счет стимуляции иными (не относящимися к имеющемуся патологическому процессу) микроорганизмами, минеральным маслом, лимфокинами, продуцируемыми Т-лимфоцитами, в меньшей степени — В-лимфоцитами.
Макрофаги активно участвуют в резорбции кости и хряща. При электронномикроскопическом исследовании на границе паннуса и суставного хряща обнаружены макрофаги, тесно связанные с частичками переваренных коллагеновьгх волокон. То же явление отмечено и при контакте макрофагов с резорбируемой костью.
Таким образом, макрофаги играют важную роль в развитии воспалительного процесса, его поддержании и хронизации и уже априорно могут рассматриваться как одна из главных «мишеней» антиревматической терапии.
Опубликовал Константин Моканов
Источник
Эффекты воспаления. Макрофаги и нейтрофилы при воспалении
Одним из первых результатов воспаления является «ограждение» области повреждения от остальных тканей. Тканевые пространства и лимфатические сосуды в воспаленной области блокируются сгустками фибрина, поэтому через небольшой промежуток времени жидкость с трудом протекает через интерстиций. Этот процесс отгораживания задерживает распространение бактерий или токсических продуктов.
Интенсивность воспалительного процесса обычно пропорциональна степени повреждения ткани. Например, когда стафилококки проникают в ткани, они выделяют смертельно опасные клеточные токсины. В результате воспаление развивается быстро, гораздо быстрее, чем сами стафилококки могут размножиться и распространиться. Следовательно, для локальной стафилококковой инфекции характерно быстрое отгораживание, предохраняющее от распространения ее по телу.
Стрептококки, напротив, не вызывают такого интенсивного локального разрушения тканей. Следовательно, процесс отгораживания развивается медленно, на протяжении многих часов, в течение которых стрептококки успевают размножиться и мигрировать. В результате стрептококки часто проявляют гораздо более выраженную тенденцию к распространению в организме и развитию смертельных состояний, чем стафилококки, несмотря на то, что стафилококки гораздо более губительны для тканей.
Тканевые макрофаги — первая «линия обороны» против инфекции. В течение нескольких минут после начала воспаления макрофаги, уже присутствующие в тканях, немедленно начинают свое фагоцитарное действие, будь то гистиоциты в подкожных тканях, альвеолярные макрофаги в легких, микроглия в мозге или др. При активации продуктами инфицирования и воспаления первым эффектом является быстрое увеличение каждого из макрофагов.
Затем многие из ранее неподвижных макрофагов отделяются от мест их прикрепления и становятся мобильными, формируя первую «линию обороны» против инфекции в течение примерно первого часа. Количество этих рано мобилизуемых макрофагов часто незначительно, но они спасают жизнь.
Вторжение нейтрофилов в воспаленную область является второй «линией обороны». В течение примерно первого часа после начала воспаления большое число нейтрофилов начинает внедряться в воспаленную область из крови. Этот процесс вызывается продуктами воспаленных тканей, которые инициируют следующие реакции: (1) изменяют внутреннюю поверхность эндотелия капилляров, способствуя прилипанию нейтрофилов к стенкам капилляров в воспаленной области.
Этот эффект, показанный на рисунке, называют маргинацией (или краевым стоянием лейкоцитов), (2) ведут к разрыхлению сцеплений между эндотелиальными клетками капилляров и небольших венул, обеспечивая появление достаточно больших отверстий для прохождения нейтрофилов путем диапедеза непосредственно из крови в тканевые пространства; (3) другие продукты воспаления вызывают хемотаксис нейтрофилов в направлении поврежденных тканей, как объяснялось ранее.
Таким образом, в течение нескольких часов после начала повреждения тканей область обильно заполняется нейтрофилами. Поскольку нейтрофилы крови — уже зрелые клетки, они готовы немедленно начать свои очистительные функции для уничтожения бактерий и удаления инородного материала.
— Также рекомендуем «Нейтрофилия. Защитные механизмы воспаления»
Оглавление темы «Клетки иммунитета. Виды иммунитета»:
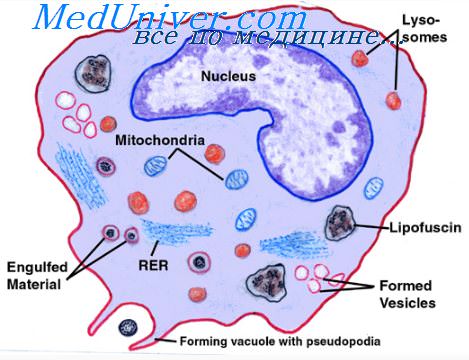
1. Ретикулоэндотелиальная система. Макрофаги в лимфатических узлах
2. Альвеолярные макрофаги в легких. Клетки Купфера печени
3. Эффекты воспаления. Макрофаги и нейтрофилы при воспалении
4. Нейтрофилия. Защитные механизмы воспаления
5. Образование гноя. Эозинофилы и базофилы
6. Лейкопения. Лейкемии и его типы
7. Врожденный иммунитет. Приобретенный или адаптивный иммунитет
8. Типы приобретенного иммунитета. Лимфоциты в приобретенном иммунитете
9. Длительность жизни белых клеток крови. Нейтрофилы и макрофаги
10. Роль лимфоцитарных клонов. Происхождение клонов лимфоцитов
Источник
Сто тридцать лет назад замечательный русский исследователь И.И. Мечников в опытах на личинках морских звезд из Мессинского пролива сделал удивительное открытие, круто изменившее не только жизнь самого будущего Нобелевского лауреата, но и перевернувшее тогдашние представления об иммунной системе.
Втыкая в прозрачное тело личинки розовый шип, ученый обнаружил, что занозу окружают и атакуют крупные амебоидные клетки. И если чужеродное тело было небольшим, эти блуждающие клетки, которые Мечников назвал фагоцитами (от греч. пожиратель), могли полностью поглотить пришельца.
Долгие годы считалось, что фагоциты выполняют в организме функции «войск быстрого реагирования». Однако исследования последних лет показали, что благодаря своей огромной функциональной пластичности эти клетки еще и «определяют погоду» многих метаболических, иммунологических и воспалительных процессов, как в норме, так и при патологии. Это делает фагоциты перспективной мишенью при разработке стратегии лечения ряда тяжелых заболеваний человека
Подвижные клетки иммунной системы – фагоциты или макрофаги, присутствуют практически во всех тканях организма. Они осуществляют активный захват, переваривание и обезвреживание чужеродных микроорганизмов (вирусов, бактерий, одноклеточных, паразитов), а также утилизацию биологического «мусора», такого как «невостребованные» и погибшие клетки (например, «стареющие» эритроциты).
В зависимости от своего микроокружения тканевые макрофаги могут выполнять и различные специализированные функции. Например, макрофаги костной ткани – остеокласты, также занимаются выведением из кости гидроксиапатита кальция. При недостаточности этой функции развивается мраморная болезнь – кость становится чрезмерно уплотненной и при этом хрупкой.
Но самым, пожалуй, удивительным свойством макрофагов оказалась их огромная пластичность, т. е. способность изменять свою транскрипционную программу («включение» тех или иных генов) и свой облик (фенотип). Следствием этой особенности является высокая разнородность клеточной популяции макрофагов, среди которых присутствуют не только «агрессивные» клетки, встающие на защиту организма-хозяина; но и клетки с «полярной» функцией, отвечающие за процессы «мирного» восстановления поврежденных тканей.
Липидные «антенны»
Своей потенциальной «многоликостью» макрофаг обязан необычной организации генетического материала – так называемому открытому хроматину. Этот не до конца изученный вариант структуры клеточного генома обеспечивает быстрое изменение уровня экспрессии (активности) генов в ответ на различные стимулы.

Выполнение макрофагом той или иной функции зависит от характера получаемых им стимулов. Если стимул будет распознан как «чужой», то происходит активация тех генов (и соответственно функций) макрофага, которые направлены на уничтожение «пришельца». Однако макрофаг могут активировать и сигнальные молекулы самого организма, которые побуждают эту иммунную клетку участвовать в организации и регуляции обмена веществ. Так, в условиях «мирного времени», т. е. при отсутствии патогена и обусловленного им воспалительного процесса, макрофаги участвуют в регуляции экспрессии генов, отвечающих за метаболизм липидов и глюкозы, дифференцировку клеток жировой ткани.
Интеграция между взаимоисключающими «мирным» и «военным» направлениями работы макрофагов осуществляется путем изменения активности рецепторов клеточного ядра, представляющих собой особую группу регуляторных белков.
Среди этих ядерных рецепторов следует особо выделить так называемые липидные сенсоры, т. е. белки, способные взаимодействовать с липидами (например, окисленными жирными кислотами или производными холестерина) (Смирнов, 2009). Нарушение работы этих чувствительных к липидам регуляторных белков в макрофагах может быть причиной системных обменных нарушений. Например, дефицит в макрофагах одного из этих ядерных рецепторов, обозначаемых как PPAR-гамма, приводит к развитию диабета 2 типа и дисбалансу липидного и углеводного обмена во всем организме.
Клеточные метаморфозы
В разнородном сообществе макрофагов на основе базовых характеристик, определяющих их принципиальные функции, выделяют три основных клеточных субпопуляции: макрофаги М1, М2 и Мox, которые участвуют, соответственно, в процессах воспаления, восстановления поврежденных тканей, а также защите организма от окислительного стресса.
«Классический» макрофаг М1 формируется из клетки-предшественника (моноцита) под действием каскада внутриклеточных сигналов, запускающихся после распознавания инфекционного агента с помощью специальных рецепторов, расположенных на поверхности клетки.
Образование «пожирателя» М1 происходит в результате мощной активации генома, сопровождаемой активацией синтеза более чем сотни белков – так называемых факторов воспаления. К ним относятся ферменты, способствующие генерации свободных радикалов кислорода; белки, привлекающие в очаг воспаления другие клетки иммунной системы, а также белки, способные разрушать оболочку бактерий; воспалительные цитокины – вещества, обладающие свойствами активировать иммунные клетки и оказывать токсическое действие на остальное клеточное окружение. В клетке активируется фагоцитоз и макрофаг начинает активно разрушать и переваривать все, что встретится на его пути (Шварц, Свистельник, 2012). Так появляется очаг воспаления.
Однако уже на начальных этапах воспалительного процесса макрофаг М1 начинает активно секретировать и противовоспалительные субстанции – низкомолекулярные липидные молекулы. Эти сигналы «второго эшелона» начинают активировать вышеупомянутые липидные сенсоры в новых «рекрутах»-моноцитах, прибывающих в очаг воспаления. Внутри клетки запускается цепь событий, в результате которых активирующий сигнал поступает на определенные регуляторные участки ДНК, усиливая экспрессию генов, отвечающих за гармонизацию обмена веществ и одновременно подавляя активность «провоспалительных» (т. е. провоцирующих воспаление) генов (Душкин, 2012).

Так в результате альтернативной активации образуются макрофаги М2, которые завершают воспалительный процесс и способствуют тканевому восстановлению. Популяцию М2 макрофагов можно, в свою очередь, разделить на группы в зависимости от их специализации: уборщики мертвых клеток; клетки, участвующие в реакции приобретенного иммунитета, а также макрофаги, секретирующие факторы, которые способствуют замещению погибших тканей соединительной тканью.
Еще одна группа макрофагов – Мох, формируется в условиях так называемого окислительного стресса, когда в тканях возрастает опасность повреждения их свободными радикалами. Например, Мох составляют около трети всех макрофагов атеросклеротической бляшки. Эти иммунные клетки не только сами устойчивы к повреждающим факторам, но и участвуют в антиоксидантной защите организма(Gui et al., 2012).
Пенистый камикадзе
Одной из самых интригующих метаморфоз макрофага является его превращение в так называемую пенистую клетку. Такие клетки были обнаружены в атеросклеротических бляшках, а свое название получили из-за специфического внешнего вида: под микроскопом они напоминали мыльную пену. По сути, пенистая клетка – это тот же макрофаг М1, но переполненный жировыми включениями, преимущественно состоящими из водонерастворимых соединений холестерина и жирных кислот.
Была высказана гипотеза, ставшая общепринятой, что пенистые клетки образуются в стенке атеросклеротических сосудов в результате неконтролируемого поглощения макрофагами липопротеинов низкой плотности, переносящих «плохой» холестерин. Однако впоследствии было обнаружено, что накопление липидов и драматическое (в десятки раз!) возрастание скорости синтеза ряда липидов в макрофагах можно спровоцировать в эксперименте только лишь одним воспалением, без всякого участия липопротеинов низкой плотности (Душкин, 2012).
Это предположение подтвердилось клиническими наблюдениями: оказалось, что превращение макрофагов в пенистую клетку происходит при разнообразных заболеваниях воспалительной природы: в суставах – при ревматоидном артрите, в жировой ткани – при диабете, в почках – при острой и хронической недостаточности, в ткани мозга – при энцефалитах. Однако понадобилось около двадцати лет исследований, чтобы понять, как и зачем макрофаг при воспалении превращается в клетку, нафаршированную липидами.
 уже в самые первые часы воспаления, вызванного определенным стимулом (например, бактерией). Макрофаги М1 и пенистые клетки начинают продуцировать провоспалительные факторы и кислородные метаболиты и активно захватывать «чужеродные» молекулы. На 1—3-е сутки воспалительного процесса пенистые клетки начинают секретировать противовоспалительные факторы, которые активируют липидные сенсоры моноцитов, мигрирующих из русла крови в очаг воспаления. Так образуются макрофаги М2. Сами пенистые клетки погибают в ходе запрограммированной клеточной смерти (апоптоза) и поглощаются макрофагами М2. Это сигнал о завершении воспаления (5-е сутки). На фото – фазово-контрастная микроскопия клеток, полученных из перитонеальной полости мышей при воспалении")
Оказалось, что активация провоспалительных сигнальных путей в М1 макрофагах приводит к «выключению» тех самых липидных сенсоров, которые в нормальных условиях контролируют и нормализуют липидный обмен (Душкин, 2012). При их «выключении» клетка и начинает накапливать липиды. При этом образующиеся липидные включения представляют собой вовсе не пассивные жировые резервуары: входящие в их состав липиды обладают способностью усиливать воспалительные сигнальные каскады. Главная цель всех этих драматических изменений – любыми средствами активировать и усилить защитную функцию макрофага, направленную на уничтожение «чужих» (Melo, Drorak, 2012).
Однако высокое содержание холестерина и жирных кислот дорого обходится пенистой клетке – они стимулируют ее гибель путем апоптоза, запрограммированной клеточной смерти. На внешней поверхности мембраны таких «обреченных» клеток обнаруживается фосфолипид фосфатидилсерин, в норме расположенный внутри клетки: появление его снаружи является своеобразным «похоронным звоном». Это сигнал «съешь меня», который воспринимают М2 макрофаги. Поглощая апоптозные пенистые клетки, они начинают активно секретировать медиаторы заключительной, восстановительной стадии воспаления.
Фармакологическая мишень
Воспаление как типовой патологический процесс и ключевое участие в нем макрофагов является, в той или иной мере, важной составляющей в первую очередь инфекционных заболеваний, вызванных различными патологическими агентами, от простейших и бактерий до вирусов: хламидиальные инфекции, туберкулез, лейшманиоз, трипаносомоз и др. Вместе с тем макрофаги, как уже упоминалось выше, играют важную, если не ведущую, роль в развитии так называемых метаболических заболеваний: атеросклероза (главного виновника сердечно-сосудистых заболеваний), диабета, нейродегенеративных заболеваний мозга (болезнь Альцгеймера и Паркинсона, последствия инсультов и черепно-мозговых травм), ревматоидного артрита, а также онкологических заболеваний.
Разработать стратегию управления этими клетками при различных заболеваниях позволили современные знания о роли липидных сенсоров в формировании различных фенотипов макрофага.
Так, оказалось, что в процессе эволюции хламидии и туберкулезные палочки научились использовать липидные сенсоры макрофагов, чтобы стимулировать не опасную для них альтернативную (в М2) активацию макрофагов. Благодаря этому поглощенная макрофагом туберкулезная бактерия может, купаясь как сыр в масле в липидных включениях, спокойно дожидаться своего освобождения, а после гибели макрофага размножаться, используя содержимое погибших клеток в качестве пищи (Melo, Drorak, 2012).
Если в этом случае использовать синтетические активаторы липидных сенсоров, которые препятствуют образованию жировых включений и, соответственно, предотвращают «пенистую» трансформацию макрофага, то можно подавить рост и понизить жизнеспособность инфекционных патогенов. По крайней мере в экспериментах на животных уже удалось в разы снизить обсемененность легких мышей туберкулезными бациллами, используя стимулятор одного из липидных сенсоров или ингибитор синтеза жирных кислот (Lugo-Villarino et al., 2012).
Еще один пример – такие болезни, как инфаркт миокарда, инсульт и гангрена нижних конечностей, опаснейшие осложнения атеросклероза, к которым приводит разрыв так называемых нестабильных атеросклеротических бляшек, сопровождаемый моментальным образованием тромба и закупоркой кровеносного сосуда.
Формированию таких нестабильных атеросклеротических бляшек и способствует макрофаг М1/пенистая клетка, который продуцирует ферменты, растворяющие коллагеновое покрытие бляшки. В этом случае наиболее эффективная стратегия лечения – превращение нестабильной бляшки в стабильную, богатую коллагеном, для чего требуется трансформировать «агрессивный» макрофаг М1 в «умиротворенный» М2.
Экспериментальные данные свидетельствуют, что подобной модификации макрофага можно добиться, подавляя в нем продукцию провоспалительных факторов. Такими свойствами обладает ряд синтетических активаторов липидных сенсоров, а также природные вещества, например, куркумин – биофлавоноид, входящий в состав корня куркумы, хорошо известной индийской пряности.
Нужно добавить, что такая трансформация макрофагов актуальна при ожирении и диабете 2 типа (большая часть макрофагов жировой ткани имеет М1 фенотип), а также при лечении нейродегенеративных заболеваний мозга. В последнем случае в мозговых тканях происходит «классическая» активация макрофагов, что приводит к повреждению нейронов и накоплению токсичных веществ. Превращение М1-агрессоров в мирных дворников М2 и Mox, уничтожающих биологический «мусор», может в ближайшее время стать ведущей стратегией лечения этих заболеваний (Walace, 2012).
С воспалением неразрывно связано и раковое перерождение клеток: например, имеются все основания считать, что 90 % опухолей в печени человека возникает как следствие перенесенных инфекционных и токсических гепатитов. Поэтому с целью профилактики раковых заболеваний необходимо контролировать популяцию М1 макрофагов.
Однако не все так просто. Так, в уже сформированной опухоли макрофаги преимущественно приобретают признаки статуса М2, который содействует выживанию, размножению и распространению самих раковых клеток. Более того, такие макрофаги начинают подавлять противораковый иммунный ответ лимфоцитов. Поэтому для лечения уже образовавшихся опухолей разрабатывается другая стратегия, основанная на стимулировании у макрофагов признаков классической М1-активации (Solinas et al., 2009).
Примером такого подхода служит технология, разработанная в новосибирском Институте клинической иммунологии СО РАМН, при которой макрофаги, полученные из крови онкобольных, культивируют в присутствии стимулятора зимозана, который накапливается в клетках. Затем макрофаги вводят в опухоль, где зимозан освобождается и начинает стимулировать классическую активацию «опухолевых» макрофагов.
, необходимо способствовать уменьшению количества «агрессивных» макрофагов М1 и увеличению «мирных» макрофагов М2. При заболеваниях же инфекционной природы и опухолях требуется, наоборот, увеличить число макрофагов-пожирателей М1")
Сегодня становится все более очевидно, что соединения, вызывающие метаморфозы макрофагов, оказывают выраженное атеропротективное, антидиабетическое, нейропротективное действие, а также защищают ткани при аутоиммунных заболеваниях и ревматоидном артрите. Однако такие препараты, имеющиеся на сегодня в арсенале практикующего врача, – фибраты и производные тиазолидона, хотя и снижают смертность при этих тяжелых заболеваниях, но при этом имеют выраженные тяжелые побочные действия.
Эти обстоятельства стимулируют химиков и фармакологов к созданию безопасных и эффективных аналогов. За рубежом – в США, Китае, Швейцарии и Израиле уже проводятся дорогостоящие клинические испытания подобных соединений синтетического и природного происхождения. Несмотря на финансовые трудности, российские, в том числе и новосибирские, исследователи также вносят свой посильный вклад в решение этой проблемы.
Так, на кафедре химии Новосибирского государственного университета было получено безопасное соединение TS-13, стимулирующее образование Мox фагоцитов, которое обладает выраженным противовоспалительным эффектом и оказывает нейропротективное действие в экспериментальной модели болезни Паркинсона (Дюбченко и др., 2006; Зенков и др., 2009).
В Новосибирском институте органической химии им. Н. Н. Ворожцова СО РАН созданы безопасные антидиабетические и противоатеросклеротические препараты, действующие сразу на несколько факторов, благодаря которым «агрессивный» макрофаг М1 превращается в «мирный» М2 (Dikalov et al., 2011). Большой интерес вызывают и растительные препараты, получаемые из винограда, черники и других растений с помощью механохимической технологии, разработанной в Институте химии твердого тела и механохимии СО РАН (Dushkin, 2010).
С помощью финансовой поддержки государства можно в самое ближайшее время создать отечественные средства для фармакологических и генетических манипуляций с макрофагами, благодаря которым появится реальная возможность превращать эти иммунные клетки из агрессивных врагов в друзей, помогающих организму сохранить или вернуть здоровье.
Литература
Душкин М. И. Макрофаг/пенистая клетка как атрибут воспаления: механизмы образования и функциональная роль // Биохимия, 2012. T. 77. C. 419—432.
Смирнов А. Н. Липидная сигнализация в контексте атерогенеза // Биохимия. 2010. Т. 75. С. 899—919.
Шварц Я. Ш., Свистельник А. В.Функциональные фенотипы макрофагов и концепция М1-М2-поляризации. Ч. 1 Провоспалительный фенотип. // Биохимия. 2012. Т. 77. С. 312—329.
Источник