
Что такое иммунный ответ при воспалении

Оглавление темы «Селезенка. Лимфатические узлы. Стадии иммунного ответа. Формы иммунного ответа. Воспаление. Механизмы, контролирующие иммунную систему.»: Стадии иммунного ответа. Формы иммунного ответа. Воспаление. Ранний защитный воспалительный ответ.Ранний защитный воспалительный ответ призван препятствовать внедрению и распространению возбудителя, по возможности быстро удалять его из организма. Ранний защитный воспалительный ответ разыгрывается в течение первых 4 сут после внедрения возбудителя. Ранний защитный ответ против внедрившихся в организм патогенных микроорганизмов обеспечивается факторами врожденного иммунитета, к которым относятся фагоцитирующие клетки крови и тканей, естественные киллеры, циркулирующие в крови белковые молекулы, обладающие защитными свойствами (компоненты системы комплемента и др.), а также межклеточные медиаторы — цитокины. Ранний воспалительный ответ стимулирует последующий специфический иммунный ответ, влияет на его форму, способствуя развитию наиболее эффективного против конкретного микроорганизма специфического иммунного ответа.
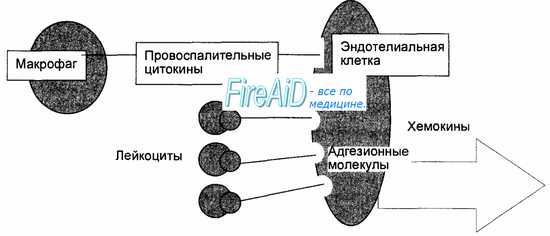
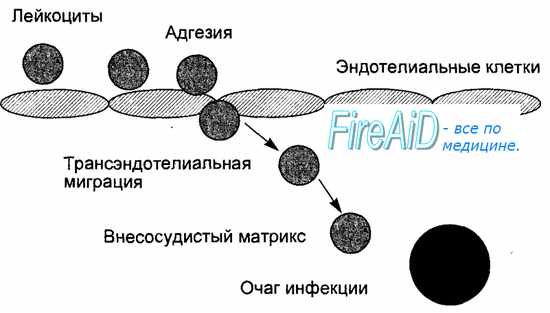
Ранний воспалительный ответ начинается с привлечения лейкоцитов из кровяного русла в очаг инфекции с последующей их активацией для удаления возбудителя (рис. 8.6). Мобилизация лейкоцитов в очаг инфекции стимулируется провоспалительными цитокинами, а опосредуется адгезионными молекулами на поверхности лейкоцитов и эндотелиальных клеток, а также молекулами хемокинов и их рецепторами. Провоспалительные цитокины (туморнекротизирующий фактор—ТНФ, интерлейкин-1 — ИЛ-1) продуцируются и секретируются макрофагами при фагоцитозе возбудителя. Под действием этих цитокинов на поверхности эндотелиальных клеток, появляются адгезионные молекулы вначале для нейтрофилов, а затем для моноцитов и лимфоцитов, ответственные за прилипание лейкоцитов к эндотелию сосудов. Кроме того, активированные цитокинами эндотелиальные клетки продуцируют хемокины, достигающие высокой концентрации на уровне очага инфекции (рис. 8.7). Этим обеспечивается прочная адгезия лейкоцитов к эндотелиальным клеткам с последующей их трансэндотелиальной миграцией и выходом в ткани в направлении очага инфекции (см. рис. 8.6). Таким образом, ранний воспалительный ответ проявляется инфильтрацией очага инфекции фагоцитирующими клетками, где эти клетки получают дополнительные сигналы активации от микробных продуктов и компонентов (липополисахарид клеточной стенки бактерий), от компонентов активированной системы комплемента и от провоспалительных цитокинов, в том числе, от гамма-интерферона, продуцируемого и секретируемого активированными естественными киллерами. Активированные фагоцитирующие клетки приобретают повышенную способность убивать захваченных микробов, что способствует удалению возбудителя. Активированные под влиянием провоспалительных цитокинов эндотелиальные клетки продуцируют молекулы вазодилататоров, под действием которых усиливается местный кровоток, повышается проницаемость сосудов для макромолекул фибриногена, который, покидая сосуды, превращается в фибрин, что способствует ограничению очага инфекции. В случае попадания в организм небольшого количества низковирулентных возбудителей ранний воспалительный ответ подавляет очаг инфекции.
Удаление из кровяного русла попавших в кровь единичных бактериальных клеток является функцией системы комплемента. Большая часть компонентов комплемента синтезируются гепатоцитами и мононуклеарными фагоцитами. Компоненты комплемента (С1, С2, СЗ, С4, С5, С6, С7, С8, С9, факторы В и D) содержатся в крови в неактивной форме. При попадании в кровяное русло бактерий на их поверхности каскад ферментативных реакций ведет к последовательной активации компонентов системы комплемента («альтернативный путь активации») с формированием мембран-атакующего комплекса (С5—С9), вызывающего лизис бактерий. В процессе активации системы комплемента накапливаются фрагменты, которые опосредуют разные биологические эффекты: привлечение лейкоцитов в очаг инфекции или воспаления (хемотаксис) — фрагмент С5а, усиление фагоцитоза (опсонизацию) — СЗb, индукцию синтеза и секреции медиаторов воспаления — С3а, С5а. (При развитии специфического иммунного ответа и появлении в кровяном русле комплексов антиген—антитело другой каскад ферментативных реакций ведет к активации системы комплемента — «классический путь активации») — Также рекомендуем «Представление антигена. Распознавание антигена. Взаимодействие Т-хелперов ( Тh1 ) с антигенпредставляющими клетками.» |
Источник
Схематическое представление первичной и вторичной иммунной реакции
Иммунный ответ — это сложная многокомпонентная, кооперативная реакция иммунной системы организма, индуцированная антигеном, уже распознанным как чужеродный, и направленная на его элиминацию. Явление иммунного ответа лежит в основе иммунитета. Иммунный ответ зависит от:
- антигена — свойства, состав, молекулярная масса, доза, кратность попадания, длительность контакта;
- состояния организма (иммунологическая реактивность);
- условий внешней среды.
Иммунный ответ различается врождённый и приобретённый (или адаптивный). Врождённый — это распознавание чужеродных раздражителей благодаря унаследованным механизмам, в то время как приобретённый иммунный ответ использует для распознавания рецепторы, число которых почти ничем не ограничено и они формируются в организме каждого человека. Приобретённый иммунный ответ может гибко реагировать на чужеродный раздражитель: если тот будет сочтён не опасным, то данный раздражитель в будущем больше не будет вызвать реакции (иммунологическая толерантность). Если же раздражитель классифицирован как опасный, последует продуктивный иммунный ответ, и чужеродные цели-раздражители будут устранены.
Иммунный ответ действует на вторжение болезнетворных микроорганизмов в организм (бактерии, вирусы, грибки, паразиты) либо патологически изменённые клетки собственного тела (злокачественная опухоль). Иммунный ответ против нормальных клеток организма может возникать, когда собственные вещества организма ошибочно воспринимаются как чужеродные (аутоиммунные заболевания). Кроме этого, безвредные вещества из окружающей среды также ошибочно могут быть классифицированы как опасные и вызывать реакцию (аллергия). С другой стороны, если патогенам удаётся избежать иммунного ответа, то это приводит к ускользанию от иммунного ответа (нем. Immunevasion).
Механизмы, которые вызывают устранение вторгнувшихся в организм патогенов в ходе иммунного ответа, чрезвычайно разнообразны. Система комплемента состоит из самоорганизующихся белковых комплексов, которые помечают агента или могут напрямую убить его. Микробицидные вещества высвобождаются клетками вне иммунной системы, также могут быть активированы разнообразные макрофаги и клетки-киллеры, а антитела начнут связываться с высокоспецифичными чужеродными структурами. Обычно несколько таких эффекторных механизмов действуют одновременно с тем, чтобы обеспечить полное удаление патогенов из организма. Тем не менее, чрезмерная реакция иммунной системы должна предотвращаться, так как это может привести к серьёзным повреждениям или даже некрозу тканей, а также летальному исходу (анафилактический или септический шок, гиперцитокинемия). Кроме того, значительная реакция против собственного тела также должна быть исключена, чтобы предотвратить аутоиммунные заболевания. Таким образом, в иммунной системе присутствует множество регуляторных механизмов для того, чтобы соблюдался баланс между защитным и вредным иммунным ответом.
Врождённый иммунный ответ[править | править код]
Во врождённом иммунном ответе участвует большое разнообразие типов клеток и переменных факторов, что в совокупности образует тесно связанную и скоординированную форму защитной системы. Это обусловлено характеристиками возбудителей иммунной реакции, которые распознаются врождёнными рецепторами — отсюда термин «врождённая иммунная реакция». Такой иммунный ответ происходит быстро и эффективно: через считанные минуты после проникновения в организм большинство болезнетворных микроорганизмов обнаруживаются и обезвреживаются, а спустя несколько часов они полностью устранены.
Приобретённый иммунный ответ[править | править код]
Посредством фагоцитоза активированные вирусом макрофаги затем «назначаются» иммунной системой для специфического или адаптивного приобретённого иммунного ответа. В свою очередь он различается на 2 вида: гуморальный и клеточный иммунный ответ.
Гуморальный иммунный ответ[править | править код]
Гуморальный иммунный ответ организма представляет собой антитела против патогенов, в основном находящихся в жидкости крови и лимфы, а также в бесклеточной плазме или в сыворотке крови. Специфические антитела, также называемые иммуноглобулинами, — это белковые молекулы, которые продуцируются и высвобождаются плазматическими клетками и действуют в кровяном и лимфатическом потоках.
Плазматические клетки являются потомками активированных B-лимфоцитов. Активация B-лимфоцита происходит путём связывания соответствующего антигена с распознающим антигены B-клеточным рецептором. Это специфические иммуноглобулины, закреплённые в клеточной мембране B-клетки.
Гиперчувствительность и аллергия[править | править код]
Гиперчувствительность (создаваемая антителами анафилаксия и клеточно-опосредованная аллергия) является приобретённым иммунным ответом с «памятью». Такой иммунный ответ может наступать в качестве контактного дерматита против химически чистых веществ после первичного или нескольких повторных контактов с аллергеном. Клеточно-опосредованным иммунным ответом может являться отторжение экзогенных трансплантированных органов.
Литература[править | править код]
- Jörg Hacker, Jürgen Heesemann. Molekulare Infektionsbiologie. — 1-е издание. — Heidelberg, Berlin: Spektrum Akademischer Verlag, 2000. — ISBN 3-86025-368-9.
- Michael T. Madigan, John M. Martinko. Brock Biology of Microorganisms. — 11-е международное издание. — Upper Saddle River, NJ., USA: Pearson Prentice Hall, 2006. — 992 с. — ISBN 978-0131443297.
- Peter F. Zipfel, Peter Kraiczy, Jens Hellwage: Das tägliche Versteckspiel: Wie Mikroorganismen der Immunabwehr entgehen. Biologie in unserer Zeit 32(6), S. 371 — 379 (2002), ISSN 0045-205X
- Diethard Baron, Jürgen Braun, Andreas Erdmann: Grüne Reihe. Genetik. Materialien S II. (Lernmaterialien)
- Charles A. Janeway, Paul Travers, Mark Walport: Immunobiology. B&T; 6. Auflage (2005) ISBN 0815341016.
Источник
Иммунокомплексный вариант воспаления
Воспаление может быть ответом на образование продуктов протеолиза компонентов комплемента при активации системы комплемента при участии иммунных комплексов.
Среди таких продуктов протеолиза особое место занимают хемоаттрактанты и анафилатоксины (С3а, С4а, С5а), которые стимулируют тучные клетки и гранулоциты к дегрануляции и секреции вазоактивных медиаторов.
Анафилатоксины индуцируют секрецию медиаторов, вызывающих быстрое повышение проницаемости сосудов.
Рецепторы для С3а и С4а экспрессированы на тучных клетках, базофилах, гладко-мышечных клетках, лимфоцитах. Рецепторы для С5а экспрессированы на тучных клетках, базофилах, нейтрофилах, моноцитах/макрофагах, эндотелиальных клетках. Экспрессия С5а-рецепторов была выявлена и на эпителиальных клетках бронхов и легких, и на гладкомышечных клетках и эндотелиальных клетках сосудов легких. При связывании анафилатоксинов со специфическими для них рецепторами на базофилах и тучных клетках индуцируется экзоцитоз гранул, содержащих вазоактивные медиаторы (гистамин и др.).
Пептид С5а обладает дополнительно активностью хемоаттрактанта для гранулоцитов, которых он заставляет мигрировать строго по градиенту его концентрации. Пептид С5а стимулирует окислительный метаболизм нейтрофилов, их дегрануляцию и адгезию к эндотелию, повышая одновременно проницаемость эндотелия. Кроме того, С5а индуцирует секрецию гистамина тучными клетками (рис. 6.4).

Рисунок 6.4. Участие продуктов активации комплемента в патогенезе иммунного воспаления. Связанные с клетками антитела или иммунные комплексы могут инициировать активацию системы комплемента по классическому пути. Образующиеся при этом анафилатоксины С3а и С5а могут непосредственно вызывать дегрануляцию тучных клеток. С5а является хемоаттрактантом для нейтрофилов и обеспечивает их приток в очаг воспаления. Эти клетки секретируют активные медиаторы и разрушающие ферменты. Медиаторы влияют на функции лейкоцитов, эндотелия и гладких мышц сосудистой стенки. Ферменты вызывают повреждение тканей и активацию системы кининов. В результате повышения проницаемости сосудов развивается местный отек. Параллельно активируется система свертывания крови, что вместе с аггрегацией тромбоцитов способствует тромбозу. Продукты повреждения тканей могут усиливать активацию системы комплемента по альтернативному пути.
Сочетание этих эффектов анафилатоксинов обеспечивает аккумуляцию клеток и белков сыворотки, характерную для острого воспаления. Основными воспалительными клетками при этом являются нейтрофилы, которые очищают очаг воспаления от возбудителей и, в то же время, могут повреждать ткани организма секретируемыми продуктами: протеазами, реактивными кислородными радикалами.
Поскольку С3а пептид является хемоаттрактантом и для моноцитов/макрофагов, то продуцируемые этими клетками провоспалительные цитокины вносят свой вклад в развитие воспаления. Показана патогенетическая роль С 5а при опосредованных лейкоцитами легочных дисфункциях при остром респираторном дистресс-синдроме, бактериальных пневмониях, кистозном фиброзе, хронических заболеваниях легких.
В формирующемся на месте отложения иммунных комплексов очаге воспаления иммунных комплексов (ИК) могут связываться с воспалительными клетками через FcR или CR1 рецепторы и индуцировать местную секрецию цитокинов и вазоактивных медиаторов, которые тоже вносят свой вклад в развитие воспаления.
Для воспаления легких, опосредованного иммунными комплексами и активацией комплемента, характерно воспалительное повреждение легочной ткани. При этом последовательность событий может быть следующей: альвеолярные макрофаги активируются при воздействии иммунных комплексов через FcR и активированными компонентами комплемента через CR1, это приводит к усиленной продукции и секреции провоспалительных цитокинов TNF-a, IL-1, которые усиливают экспрессию молекул ICAM-1 и Е-селектина на сосудистом эндотелии, к которому прикрепляются нейтрофилы, и активируются при участии IL-8 и PA F из эндотелиальных клеток.
Их трансмиграция в альвеолы индуцируется С5а, который генерируется в альвеолах при взаимодействии иммунных комплексов с системой комплемента. Рекрутирование гранулоцитов из сосудов идет при участии интегринов LFA-1 и Мас-1. Активация альвеолярных макрофагов с усилением секреции TNF-a приводит к повышению экспрессии адгезионных молекул ICAM-1 на альвеолярном эпителии.
К этим адгезионным молекулам прикрепляются клетки внутри альвеол: макрофаги и нейтрофилы, которые активируются, секретируют повреждающие цитокины и ферменты. Воспалительное повреждение легких при таком типе воспаления опосредуется рекрутированными нейтрофилами.
На модели воспалительного повреждения легких после депозиции иммунных комплексов было показано, что природным регулятором воспаления служит цитокин IL-10, который способен супрессировать продукцию TNF-a, экспрессию адгезионных молекул ICAM-1 и рекрутирование нейтрофилов в очаг. За счет всех этих механизмов IL-10 уменьшает степень повреждения легких. Продукция IL-10 в легких является конститутивной, она усиливается в первые часы развития иммунокомплексного воспаления.
Иммунное воспаление — гиперчувствительность замедленного типа
Иммунное воспаление — гиперчувствительность замедленного типа (ГЗТ) представляет собой эффекторную фазу специфического клеточного иммунного ответа и включает следующие события: активацию цитокинами сосудистого эндотелия, рекрутирование моноцитов и лимфоцитов из кровяного русла и тканей в очаг ГЗТ, активацию функций макрофагов лимфокинами в очаге гиперчувствительности замедленного типа, элиминацию причинного антигена путем очищения очага ГЗТ от возбудителей и/или повреждение тканей секретируемыми продуктами активированных макрофагов и лимфоцитов. Основными участниками иммунного воспаления являются: моноциты/макрофаги, Т-лимфоциты (ТН1) и эндотелиальные клетки (рис. 6.5).
Рисунок 6.5. Реакция гиперчувствительности замедленного типа (ГЗТ). В верхней левой части рисунка — стадия сенсибилизации при первой встрече с антигеном включает: представление антигена антиген-представляющей клеткой (АРС) Т-лимфоциту (ТН) с последующей его активацией, пролиферацией, дифференцировкой в ТН1, продукцией соответствующих цитокинов. В правой нижней части рисунка — стадия ответа на повторную встречу с тем же антигеном включает: представление антигена и его распознавание ТН1, что ведет к его активации, секреции цитокинов и активации макрофагов.
В процессе иммунного воспаления ведущую роль играют следующие цитокины: IFNy, TNF-a, TNF-в, IL-1, IL-6. В реакциях гиперчувствительности замедленного типа различают острую фазу и стадию хронического воспаления. Острая фаза по своим проявлениям сходна с ранним воспалительным неспецифическим ответом, но отличается тем, что макрофаги исходно активируются не микробными продуктами, а IFNy и другими цитокинами (MIF, GM-CSF). Продукты активированных Т-лимфоцитов IL-3, GM-CSF стимулируют и продукцию моноцитов, и их рекрутирование из кровяного русла (TNF-a, TNF-в, МСР). В результате на месте очага иммунного воспаления формируется мононуклеарный инфильтрат.
В стадии хронического воспаления те же провоспалительные цитокины (IL-1, IL-6, TNF-a) стимулируют пролиферацию фибробластов и синтез коллагена как непосредственно, так и через индукцию каскада других цитокинов: PDGF, TGFp, FGF, которые в совокупности еще усиливают ангиогенез.
Сочетанный эффект перечисленных медленно действующих цитокинов и ростовых факторов при длительной неконтролируемой активации макрофагов в очаге хронического иммунного воспаления ведет к замещению тканей органов (в том числе легких) фиброзной тканью. Фиброз, как правило, сопутствует хроническому иммунному воспалению, приходящему на смену неэффективному острому воспалению, которое не привело к элиминации причинного антигена.
При хроническом течении воспаления с персистенцией сдвигов сывороточных белков, описанных выше в связи с ранним воспалительным ответом, повышенный уровень сывороточного амилоида А может вести к его отложению в интерстициальной ткани в форме фибрилл. Развивается амилоидоз, нарушающий жизненно важные функции.
При ГЗТ активированные макрофаги постепенно претерпевают ряд изменений: увеличиваются в размерах, приобретают морфологию «эпителиоидных» клеток или сливаются, образуя многоядерные гигантские клетки. Такие активированные видоизмененные макрофаги собираются в конгломераты вокруг антиген-содержащих частиц или клеток. Образуется узел воспалительной ткани — гранулема. Гранулема — это характерный ответ в виде хронической формы ГЗТ на длительно персистирующую в ткани микробную инфекцию, например, при туберкулезе или микозах, которые препятствуют распространению инфекции. Исходом гранулемы может быть деструкция ткани вплоть до некроза с последующим фиброзом.
Проникающий через респираторный тракт Cryptococcus neoformans в случаях недостаточно эффективного раннего воспалительного ответа не вычищается из ткани легкого и вызывает персистирующую хроническую инфекцию. В этих случаях защитную роль берет на себя Т-клеточный ответ, эффективность которого во многом зависит от вирулентности возбудителя. Высоковирулентные штаммы гриба вырабатывают меланин, который является скавенджером для свободных радикалов и тем самым защищает возбудителя от антимикробного действия супероксидных и нитроксидных радикалов.
При высоком уровне продукции меланина возбудитель индуцирует минимальный воспалительный ответ в легких, отсроченный и неэффективный. В этом случае снижена продукция TNF-a альвеолярными макрофагами и снижен про-лиферативный ответ Т-лимфоцитов. Дефектность клеточного иммунного ответа на меланин-продуцирующие штаммы гриба проявляется низким содержанием в легких СD4+Т-клеток, минимальным рекрутированием воспалительных клеток, сниженной активацией макрофагов, сниженным очищением легких, повышенной диссеминацией гриба в центральную нервную систему, минимальной выраженностью обоих типов ответа: ТН1 и ТН2.
В генезе индуцированной туберкулезным антигеном в ткани легкого гранулемы ведущую роль играют IFNy и TNF-a. Эти два цитокина обеспечивают максимальную выраженность местного воспаления за счет усиления экспрессии адгезионных молекул и хемокинов, необходимых для рекрутирования моноцитов/макрофагов в очаг воспаления. К тому же IFNy активирует функции макрофагов и способствует дифференцировке ТН1 лимфоцитов в региональных лимфоузлах, а также может усиливать адгезию лимфоцитов к эндотелию при их рекрутировании.
Активированные макрофаги продуцируют не только провоспалительные цитокины, но и противовоспалительные, в том числе IL-10. Этот цитокин ингибирует генерацию ТН1 лимфоцитов в региональных лимфоузлах, ингибирует их активность, препятствует формированию гранулемы и тем самым способствует диссеминации инфекции.
Результатом ГЗТ в респираторном тракте может быть развитие эксудативного воспаления и аккумуляция лейкоцитов в просвете дыхательных путей. Экссудация — выход плазмы через ткани дыхательных путей в просвет — происходит в два этапа. Сначала белки плазмы выходят из мелких сосудов через эндотелий в интерстициальную ткань. Трахео-бронхиальные микрососуды образуют густую сеть в подслизистой и проявляют очень высокую чувствительность к действию провоспалительных медиаторов.
Далее жидкость преодолевает слой эпителия и поступает в просвет дыхательных путей, откуда она может удаляться механизмами очищения. Эффекторная фаза легочной реакции ГЗТ, как правило, достигает максимума через 24 часа после повторной встречи с антигеном и проявляется накоплением в просвете жидкости, гранулоцитов и моноцитов, рекрутированных из кровяного русла.
При системной гранулематозной болезни легких — саркоидозе иммуно-гистологическими исследованиями показано совместное отложение в гранулемах фибрина и цитокина IL-1в. Высказано предположение, что в очаге хронического воспаления, где условия способствуют коагуляции и аккумуляции фибрина, взаимодействие между мононуклеарами и фибриновым матриксом ведет к продукции IL-1в.
Федосеев Г.Б.
Опубликовал Константин Моканов
Источник